
第6章 第五章歷史上的窮與富
很大一部分人類歷史允滿了窮富之間不平等的鬥爭:具有農民力量的民族與不具有農民力量的民族之間的鬥爭,或不同時期獲得農民力量的民族之間的鬥爭。糧食生產在地球上的廣大地區過去沒有出現過,這並不令人奇怪,由於生態原因,糧食生產在這些地區現在仍然難以出現或不可能出現。例如,在史前期的北美洲北極地區,無論農業或畜牧業都沒有出現過,而在歐亞大陸北極地區出現的唯一糧食生產要素是放牧馴鹿。在遠離灌溉水源的沙漠地區也不可能自發地出現糧食生產,如澳大利亞中部和美國西部的一些地方。
迫切需要說明的,反倒是何以在某些生態條件十分適宜的地區在現代以前一直直未能出現糧食生產,而在今天卻成了世界上一些最富足的農牧中心。最為令人費解的一些地區,是加利福尼亞和美國太平洋沿岸其他—些州、阿根廷的無樹大草原、澳大利亞西南部和東南部以及南非好望角地區的很大部分。這些地區的土著族群在歐洲移民來到時還仍然過著狩獵採集生活。如果我們考察一下公元前4000年的世界,即糧食生產在其最早發源地出現後幾千年的世界,我們可能也會對其他幾個現代糧倉當時竟未出現糧食生產而感到驚異。這些盛產穀物的地區包括:美國其餘所有的地區、英國、法國很大—部分地區、印度尼西亞以及非洲赤道以南的整個地區。如果我們對糧食生產追本溯源,它的最早發源地會再次使我們感到驚異。這些地方已完全不是現代糧倉,它們包括一些在今天被列為有點乾旱或生態退化的地區:伊拉克和伊朗、墨西哥、安第斯山脈、中國的部分地區以及非洲的。為什麼糧食生產首先在看似相當貧瘠的土地上形成,只是到後來才在今天最肥沃的農田和牧場發展起來?
在糧食生產賴以出現的方式方面的地理差異也同樣令人費解。在有些地方,它是獨立發展起來的,這是當地人馴化當地動植物的結果。而在其他大多數地方,則是把別的地方已經馴化的作物和牲口加以引進。既然這些原來不是獨立發展糧食生產的地區在引進馴化動植物後立刻變得適宜於史前的糧食生產,那麼這些地區的各個族群為什麼在沒有外來幫助的情況下,通過馴化當地的動植物而成為農民和牧人呢?
在的確獨立出現糧食生產的這些地區中,為什麼出現的時間差別如此之大——例如,在東亞要比在美國東部早幾千年,而在澳大利亞東部卻又從來沒有出現過?在史前時代引進糧食生產的這些地區中,為什麼引進的時間差別也如此之大——例如,在歐洲西南部要比在美國西南部早幾千年?另外,在引進糧食生產的這些地區中,為什麼在有些地區(如美國西南部)當地的狩獵採集族群採納了鄰近族群的作物和牲口而最後成為農民,而在另一些地區(如印度尼西亞和非洲赤道以南的許多地方)引進糧食生產卻引起了—場災難,使外來的糧食生產者取代了該地區原來的狩獵採集族群呢?所有這些問題都涉及不同的發展階段,而正是這些不同的發展階段決定了哪些民族成了歷史上的貧窮民族,哪些民族成了歷史上的富有民族。
在我們能夠指望回答這些問題之前,我們需要弄清楚怎樣去確定糧食生產的發源地及其出現的時間,以及某一特定作物或動物最早得到馴化的地點和時間。最明確的證據來自對一些考古遺址中出土的動植物殘骸所作的鑑定。大多數馴化的動植物物種在形態上同它們的野生祖先是不同的:例如,馴化的牛和羊形體較小,馴化的雞和蘋果形體較大,馴化的豌豆種皮較薄也較光滑,馴化的山羊角長成螺旋形而不是短彎刀狀。因此,如果能在一處有年代可考的考古遺址認出馴化動植物的殘骸,那就是有了強有力的證據,說明被時被地已有了糧食生產,而如果在某個遺址僅僅發現了野生物種,那就不能證明已有了糧食生產,而只能證明與狩獵採集生活相吻合。當然,糧食生產者,尤其是初期的糧食生產者,在繼續採集某些野生植物和獵捕野獸,這樣,他們遺址中的殘餘食物常常不但包括馴化的物種,而日也包括野生的物種。
考古學家們用碳—14年代測定法來測定遺址中的含碳物質,從而確定糧食生產的年代。這種測定法所依據的原理是這樣的:碳是生命的無所不在的基礎材料,它的成分中有很少量的放射性碳—14,而碳—14會衰變為非放射性同位素氮—14。宇宙射線不斷地在大氣中生成碳—14。植物吸收大氣中的碳,其中碳—14和普遍存在的同位素碳—12保持著—種已知的幾乎不變的比例(約1與100萬之比)。植物中的碳接下去構成了吃這些植物的食草動物的軀體,也構成了吃這些食草動物的食肉動物的軀體。不過,這些植物或動物一旦死去,它們體內碳—14含量的一半每隔5700年衰變為碳—12,直到大約4萬年後,碳—14含量變得很低而很難測出,也很難把它同受到少量的含有碳—14的現代材料的污染區別開來。因此,從考古遺址出土的材料的年代可以根據該材料內的碳—14與碳—12的比例計算出來。
放射性碳受到許多技術問題的困擾,其中兩個問題值得在這裡提—提。一個問題是:碳—14年代測定法在20世紀80年代前需要比較多的碳(幾克),比小小的種子或骨頭里碳的含量多得多。因此,科學家們常常不得不依靠測定在同一遺址附近找到的材料的年代,而這個材料被認為是與殘存的食物“有聯繫”的——就是說,是被留下食物的人同時棄置的。通常選擇的“有聯繫”的材料是燒過的木炭。
但是,考古遺址並不總是把所有同日棄置的材料巧妙密封起來的時間容器。在不同時間棄置的材料可能會混雜在一起,因為蠕蟲、囓齒目動物和其他作用力把地層給攪亂。燃燒過的木炭碎屑最後可能因此而靠近了某個死去的並在幾千年中或早或晚被吃掉的植物或動物。今天,考古學家們越來越多地用一種叫做加速質譜分析法的新技術來解決這個問題,這種新技術使碳—14年代測定法測得極小的樣本的年代,從而使人們可以直接地測得—粒小小的種子、一塊小小的骨片或其他食物殘渣的年代。近年來用碳—14年代測定法測得的年代,有的是根據這種新的直接方法(它們也有其自身的問題),有的是根據舊的間接方法。但在有些情況下,人們發現用這兩種方法測得的年代存在著巨大的差異。在由此而產生的仍未解決的爭論中,就本書的論題而言,最重要的也許是有關糧食生產在美洲出現的年代問題:20世紀60年代和70年代的間接方法測得的年代是遠在公元前7萬年,而較近的直接方法測得的年代則不早於公元前3500年。
碳—14年代測定法的第二個問題是:大氣中碳—14與碳—12的比例事實上並不是嚴格不變的,而是隨著時間上下波動的,因此,從某種不變的比例這種假定出發去計算碳—14年代測定法測得的年代經常會產生一些小小的錯誤。確定關於過去每個年代錯誤的程度,原則上可以藉助古老樹木記錄下的年輪,因為只要數一數這些年輪,就可得到每個年輪在過去的絕對日曆年代,然後再對用這種方法測定年代的木炭樣本加以分析,來確定其中碳—14與碳—12的比例。這樣,就可以對用碳—14年代測定法實測到的年代加以校正,來估計大氣中碳比例的波動情況。這樣校正的結果是:對從表面上看(即未經校正的)其年代介於公元前約1000年至6000年之間的一些材料來說,精確的(經過校正的)年代要早幾百年或1000年。近來又有人用一種交替法開始對一些年代稍早的樣本進行校正,這種方法依據的是另一種放射性衰變法,它所得出的結論是,表面上看年代約為公元前9000年的樣本的實際年代是公元前11000年左右。
考古學家們常常把經過校正的和未經過校正的年代加以區分,其方法就是對前者用大寫英文字母來寫,對後者用小寫英文字母來寫(例如,分別為3000B.C.和3000b.c.)。然而,考古文獻在這方面可能很混亂,因為許多書和論文在報告未經校正的年代時都寫作BC,而未能提到這些年代實際上是未經校正的。我在本書中所報導的關於過去15000年中一些事件的年代都是經過校正的年代。這就是為什麼讀者會注意到關於早期糧食生產問題本書中的一些年代與從某些標準參考書引用的年代存在著差異的原因。
一且人們辨認出馴化動植物的古代遺存並確定其年代,那麼人們怎樣來確定是否這個植物或動物實際上就是在這遺址附近馴化的,而不是在別處馴化,後來才傳到這個遺址來的?一個方法就是研究一下這個作物或動物的野生祖先的地理分佈圖,並推斷出野生祖先出現的地方必定就是發生過馴化的地方。例如,從地中海和埃塞俄比亞往東到印度,傳統的農民普遍種植鷹嘴豆,今天世界上鷹嘴豆的80%都是印度生產的。因此,人們可能會誤以為鷹嘴豆是在印度馴化的。但結果表明,鷹嘴豆的野生祖先只出現在土耳其的東南部。鷹嘴豆實際上是在那裡馴化的,這個解釋得到了這樣一個事實的證明,即在新石器遺址中有關可能是馴化的鷹嘴豆的最古老的發現來自土耳其東南部和敘利亞北部鄰近地區,其年代為公元前8000年左右;直到5000多年後,關於鷹嘴豆的考古證據才在印度次大陸出現。
確定某個作物或動物的馴化地點的第二個方法,是在地圖上標出每個地區馴化物種首次出現的年代。出現年代最早的地點也許就是馴化最早的地點——而如果野生物種的祖先也在那裡出現,如果它們在其他地點首次出現的年代隨著與推定的最早馴化地點距離的增加而漸次提高,從而表明馴化物種在向其他那些地點傳播,情況就尤其如此。例如,已知最早的人工栽培的二粒小麥在公元前8500年左右出現在新月沃地。其後不久,這個作物逐步向西傳播,在公元前6500年左右到達希臘,在公元前5000左右到達德國。這些年代表明二粒小麥是在新月沃地馴化的。這一結論可以用以下事實來證明:二粒小麥的野生祖先的分佈只限於從以色列到伊朗西部和土耳其這一“地區。
然而,在許多情況下,如果同樣的植物或動物是在不同的地點獨立馴化的,那麼就會出現一些複雜的情況。只要分折一下由此產生的不同地區的相同作物或動物標本在形態、遺傳或染色體方面的差異,就常常可以發現這些情況。例如,印度馴化牛中的瘤牛品種具有歐亞大陸西部牛的品種所沒有的肉峰。遺傳分析表明,現代印度牛的品種和歐亞大陸西部牛的品種在幾十萬年前就已分化了,比任何地方任何動物馴化的時間都早很多。就是說,在過去1萬年中,牛就已在印度和歐亞大陸西部獨立地馴化了,而它們原來都是在幾十萬年以前就已分化的印度和歐亞大陸西部野牛的亞種。
現在,讓我們再回到我們原先的關於糧食生產的出現這個問題上來。在世界上的不同地區,糧食生產是在何處、何時和如何發展起來的呢?
一個極端情況是:有些地區的糧食生產完全是獨立出現的,在其他地區的任何作物或動物來到之前,許多本土作物(在有些情況下還有動物)就已馴化了。目前能夠舉出詳細而又令人信服的證據的這樣的地區只有5個:西南亞,亦稱近東或新月沃地;中國;中美洲(該詞用來指墨西哥的中部和南部以及中美洲的毗連地區);南美洲的安第斯山脈地區,可能還有亞馬孫河流域的毗連地區;以及美國東部(圖5.1)。在這些糧食生產中心中,有些中心或所有中心可能實際上包含了附近的幾個或多或少獨立出現糧食生產的中心,如中國北方的黃河流域和中國南部的長江流域。
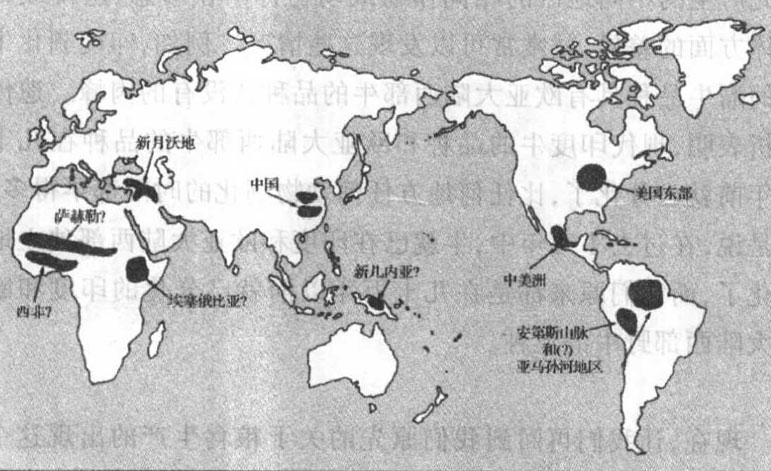
圖5.1 糧食生產發源中心
除了這5個確然無疑出現糧食生產的地區外,另外還有4個地區——非洲的薩赫勒地帶、熱帶西非、埃塞俄比亞和新幾內亞——是爭取這一榮譽稱號的候補地區。然而,每一個地區都有某種不確定之處。雖然在非洲撒哈拉沙漠南沿的薩赫勒地帶毫無疑問已有本地野生植物的馴化,但那裡牛的放牧可能在農業出現前就已開始了,目前尚不能肯定的是:這些牛是獨立馴化的薩赫勒牛,或者本來就是新月沃地飼養的牛,它們的引進引發了當地植物的馴化。同樣仍然不能肯定的是,這些薩赫勒作物的引進是否接著又在熱帶西非引發了當地人對本地野生植物的無庸置疑的馴化,而西南亞作物的引進是否就是在埃塞俄比亞引發當地人馴化本地野生植物的原因。至於新幾內亞,那裡的考古研究提供的證據表明,在任何毗連地區出現糧食生產之前很久,那裡就已有早期的農業,但那裡種植什麼作物卻一直沒有得到明確的認定。
表5.1:為在本地馴化的那些地區和其他地區扼要地列出了一些雖著名的作物或動物以及已知的最早的馴化年代。在9個獨立發展糧食生產的候補地區中,西南亞是植物馴化(公元前8500年左右)和動物馴化(公元前8000年左右)有最早的明確年代的地區;同時對於早期的糧食生產來說,它顯然也是具有最多的用碳—14測定的準確年代的地區。中國發展糧食生產的年代幾乎同西南亞一樣早,而在美國東部則顯然晚了差不多6000年。就其他6個候補地區而言,最早的得到充分證明的年代沒有超過西南亞的年代,但在這其他的6個地區由於能夠有把握確定其年代的遺址太少,我們無法肯定它們真的落後於西南亞以及(如果真的落後的話)落後多少。
下一批地區包括一些至少馴化了兩三種本地植物或動物的地區,但這些地區的糧食生產主要依靠在別處馴化的作物和動物。可以把這些引進的馴化動植物看作是“祖代”作物和動物,因為它們創立了本地的糧食生產。祖代馴化動植物的引進使本地人過著定居的生活,從而增加了野生植物演化為本地作物的可能性,這些野生植物本來是他們採集後帶回家偶然種下的,而到後來就是有意種植了。
在三四個這樣的地區,引進的祖代動植物來自西南亞。其中一個地區是歐洲的西部和中部,那裡的糧食生產是在公元前6000年和3500年之間隨著西南亞作物和動物的引進而出現的,但至少有一種植物(罌粟,可能還有燕麥和其他植物)當時是在本地馴化的。野生罌粟只生長在地中海西部沿岸地區。歐洲東部和西南亞最早的農業社會的發掘遺址中沒有發現罌粟的種子;它們的首次出現是在歐洲西部的一些早期農村遺址。與此形成對照的是,在歐洲西部卻沒有發現西南亞大多數作物和動物的野生祖先。因此,糧食生產不是在歐洲西部獨立發展起來的,這看來是很清楚的。相反,那裡的糧食生產是由於引進了西南亞的馴化動植物而引發的。由此而產生的歐洲西部農業社會馴化了罌粟,隨後罌粟就作為一種作物向東傳播。
還有一個地區,那裡由本地對動植物進行馴化,似乎是在引進西南亞的祖代作物後開始的。這個地區就是印度次大陸的印度河河谷地區。那裡的農業社會出現在公元前的第七個1000年中,它們利用的小麥、大麥和其他作物,是先前在新月沃地馴化的,然後顯然再通過伊朗傳播到印度河河谷。只是到了後來,由印度次大陸土生物種馴化的動植物,如瘤牛和芝麻,才在印度河河谷的農業社會出現。同樣,在埃及,糧食生產也是在公元前6000年隨著西南亞作物的引進而開始的。埃及人當時馴化了西克莫無花果和一種叫做鐵荸薺的植物。
同樣的模式大概也適用於埃塞俄比亞,那裡種植小麥、大麥和其他西南亞作物已有很長的歷史。為了得到作物,埃塞俄比亞人也馴化了許多在本地得到的物種,這些作物中的大多數仍然只有埃塞俄比亞才有,但其中的一種(咖啡豆)現在已傳播到全世界。然而,埃塞俄比亞人馴化這些本地植物是在西南亞馴化物種引進之前還是在引進之後,這仍然無從知曉。
在依靠從別處引進祖代作物來發展糧食生產的這些地區和其他地區,當地的狩獵採集族群是否從鄰近的農業族群那裡採納了那些祖代作物,從而使他們自己也成了農民?或者,這一攬子祖代作物竟是由入侵的農民帶來,從而使他們能夠在當地以更快的速度繁衍,並殺死、趕走或在人數上超過本地的獵人?
在埃及,似乎有可能發生前一種情況:本地的狩獵採集族群原來都是以野生動植物為食,現在又有了西南亞的馴化動植物和農牧技術,於是就逐步停止吃野生食物。這就是說,使糧食生產得以在埃及開始的是外來的作物和動物,而不是外來族群。在歐洲大西洋沿岸地區,情況也可能如此,因為那裡的狩豬採集族群在許多世紀中顯然採納了西南亞的綿羊和穀物。在南非的好望角地區,以狩獵採集為生的科伊族人,由於從遙遠的非洲北部(歸根到底還是從西南亞)得到了綿羊和牛而成為牧人(而不是農民)。同樣,美國西南部的以狩獵採集為生的印第安人,由於獲得了墨西哥的作物而成為農民。在這4個地區,糧食生產的開始幾乎沒有或根本沒有提供任何說明當地動植物馴化的證據,也幾乎沒有或根本沒有提供任何說明人口更替的證據。
另一個極端情況是:有些地區的糧食生產毫無疑問不但是從外來作物和動物的引進開始的,而且也是從外來人的突然到來開始的。我們之所以能如此肯定,是因為外來人的到來在現代也發生過,而且也與有文化的歐洲人直接有關,這些歐洲人在許多書中對所發生的事都有過描述。上面說的這些地區包括加利福尼亞、北美洲西北部太平洋沿岸、阿根廷的無樹大草原、澳大利亞和西伯利亞。直到最近幾個世紀,這些地區仍然為狩獵採集族群所佔有——在前3個地區是美洲土著,在後兩個地區是澳大利亞土著或西伯利亞土著。這些以狩獵採集為生的人遭到了陸續來到的歐洲農民和牧人的殺害、疾病的感染、驅逐、或大規模的更替。這些農民和牧人帶來了他們自己的作物,所以在來到後沒有對當地的任何野生物種進行馴化(澳大利亞的堅果樹例外)。在南非的好望角地區,陸續來到的歐洲人不但發現了科伊族中以狩獵採集為生的人,也發現了科伊族中只有馴化動物而沒有作物的牧人。結果仍然是:靠外來作物來開始農業,不馴化本地動物,以及現代人口的大規模更替。
最後,依靠外來作物來突然開始糧食生產和突然發生大規模的人口更替,這同一模式在史前時代的許多地區似乎多次出現過。由於缺乏文字記載,關於史前人口更替的證據必須從考古記錄中去尋找,或者根據語言學的證據來加以推斷。得到最充分證明的一些事例表明,人口更替現象毫無疑問是存在的,因為新來乍到的糧食生產者在骨骼方面同被他們更替的以狩獵採集為生的人有著顯著的差異,同時也因為這些糧食生產者不但引進了作物和動物,也引進了陶器。以後的幾章將對兩個最明顯的例子加以描述:南島人從華南向菲律賓和印度尼西亞的擴張(第十七章)和班圖人在非洲赤道以南地區的擴張(第十九章)。
東南歐和中歐使我們看到了一幅類似的圖景,即糧食生產(依靠西南亞的作物和動物)和製陶的突然開始。這種突然的開始大概也與古希臘人和日草曼人被現代希臘人和日耳曼人所更替直接有關,就像在菲律賓、印度尼西亞和非洲赤道以南地區舊有的人讓位於新來的人一樣。然而,原來的以狩獵採集為生的人和更替他們的農民在骨骼方面的差異,在歐洲不像在菲律賓、印度尼西亞和非洲赤道以南地區那樣顯著。因此,在歐洲人口更替的例子也就不那麼有說服力或不那麼直接。
總之,世界上只有幾個地區發展了糧食生產,而且這些地區發展糧食生產的時間也差異甚大。一些鄰近地區的狩獵採集族群從這些核心地區學會了糧食生產,而其他一些鄰近地區的族群則被來自這些核心地區的糧食生產者所更替了——更替的時間仍然差異甚大。最後,有些族群雖然生活在一些生態條件適於糧食生產產的地區,但他們在史前期既沒有發展出農業,也沒有學會農業;他們始終以狩獵採集為生,直到現代世界最後將他們淘汰。在糧食生產上具有領先優勢的那些地區裡的族群,因而在通往槍砲、病菌和鋼鐵的道路上也取得了領先的優勢。其結果就是富有社會與貧窮社會之間一系列的長期衝突。
我們怎樣來解釋糧食生產的開始在時間和模式上的地理差異呢?這個問題是關於史前史的最重要的問題之一,它將成為下面五章討論的主題。
註釋:
圖5.1 糧食生產發源中心
註釋: